Օպտիմալ կենսաբազմազանության սկզբունքը: ինչու է բնությանը անհրաժեշտ բազմազանություն և ինչպես է այն հասնում դրան
Կենսաբազմազանությունը հարմարվողականություն է: Կենսաբանական համակարգի ներքին բազմազանությունը (այսինքն՝ բաղադրիչների բազմազանությունը) հարմարվողական հատկանիշ է՝ կապված դրա արդյունավետության, առաձգականության և կենսունակության հետ: Օպտիմալ բազմազանություն ունեցող կենսահամակարգերն ամենաարդյունավետն ու կենսունակն են: Օպտիմալ բազմազանությունից ցանկացած շեղում նվազեցնում է դրանց արդյունավետությունն ու կենսունակությունը կամ նույնիսկ ի վերջո ոչնչացնում դրանք:
Օպտիմալ կենսաբազմազանությունը հաստատվում է կյանքի տարբեր հիերարխիկ մակարդակների փոխազդեցության ժամանակ: Մեր տեսական մոդելները (տե՛ս ստորև բերված հղումները) ուսումնասիրել են կյանքի հիերարխիայի միայն մի փոքր հատված, այն է՝ դրանցից բաղկացած պոպուլյացիաներն ու համայնքները, այսինքն՝ ներբնակչությունը և տեսակների բազմազանությունը: Հետագա մտածողությունը գալիս է այս մոդելներից: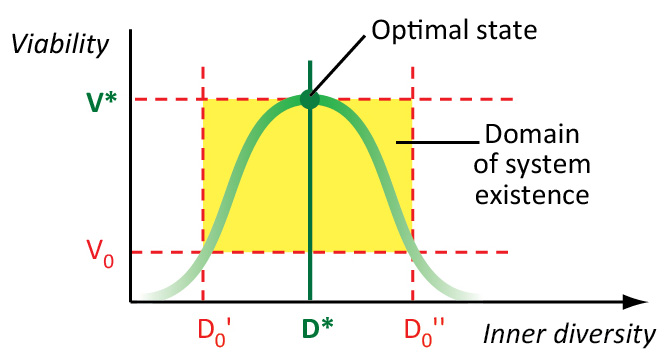
Բազմազանության օպտիմալ արժեքը (D*) համապատասխանում է կենսահամակարգի առավելագույն կենսունակությանը (V*): Վ0 - կենսունակության կրիտիկական արժեքը. Դ0 - բազմազանության կրիտիկական արժեք:
Բնակչության և համայնքների մակարդակների հակառակ արձագանքը` նույն մետաղադրամի երկու կողմերը: Ներբնակչության և տեսակների բազմազանության օպտիմալ արժեքները կախված են շրջակա միջավայրի պարամետրերից (առկա ռեսուրսների քանակից և կայունության աստիճանից) հակառակ ռեժիմում։ Ներբնակչության օպտիմալ բազմազանությունը նվազում է ավելի կայուն պայմաններում, մինչդեռ տեսակների օպտիմալ հարստությունը մեծանում է: Տեսակների օպտիմալ հարստությունը նույնպես աճում է ավելի «հարուստ» պայմաններում, իսկ ներբնակչության օպտիմալ բազմազանությունը կախված չէ ռեսուրսների քանակից: Ներբնակչությունը և տեսակների բազմազանությունը ներկայացնում են կենսաբանական հիերարխիայի երկու անբաժանելի ասպեկտներ, և դրանց ռեակցիայի տարբերությունը ցույց է տալիս հատուկ դերեր հարմարվողականության գործընթացում:
Ներքևից վեր. բաղադրիչները սահմանում են բազմազանությունը վերին մակարդակում: Տեսակների բազմազանության օպտիմալ արժեքները կախված են պոպուլյացիայի հետևյալ պարամետրերից.
- անհատների էկոլոգիական հանդուրժողականության գոտու լայնությունը.
- բնակչության աճի և մահացության առավելագույն մակարդակը.
- ֆենոտիպերի հիմնական ռեսուրսային ծախսերը և հանդուրժողականության գոտու ընդլայնման արժեքը:
Այս պարամետրերից յուրաքանչյուրի շարունակական բարելավումը հանգեցնում է օպտիմալ բազմազանության նույն փոփոխությունների, ինչպես շրջակա միջավայրի կայունացումը: Այսպիսով, պոպուլյացիաներն ունեն շրջակա միջավայրի անկայունությունը փոխհատուցելու տարբեր եղանակներ՝ ավելացնելով բնակչության աճի տեմպերը, նվազեցնել մահացությունը, ընդլայնել անհատական հանդուրժողականության գոտիները, նվազեցնելով ֆենոտիպերի ռեսուրսների ծախսերը: Այս մեխանիզմները կարող են աշխատել պոպուլյացիայի մեջ՝ շնորհիվ նրա հարմարվողական կարողությունների, և համայնքի ներսում՝ տեսակների կազմի փոփոխության պատճառով, օրինակ՝ փոխվելով K- և r-ռազմավարների և մասնագետների և գեներալիստների միջև:
Վերոնշյալ պարամետրերից որևէ մեկի շարունակական բարելավումը մյուսների անփոփոխ մնալով կարող է մեկնաբանվել որպես օրգանիզմների էվոլյուցիոն մակարդակի բարձրացում: Այսպիսով, վերջինը կարող է հանգեցնել ներբնակչության օպտիմալ բազմազանության նվազմանը և տեսակների օպտիմալ բազմազանության ավելացմանը:
Օպտիմալ կենսաբազմազանության ակնկալվող օրինաչափություններ. Ներբնակչության բազմազանությունը, այլ հավասար պայմանների դեպքում, կարելի է համարել էկոլոգիական խորշի լայնության վստահելի: Այսպիսով, մենք կարող ենք կանխատեսել համայնքների կենսաբազմազանության օպտիմալ կառուցվածքը. Չխաթարված գագաթնակետային համայնքները և նրանց բաղկացուցիչ բնակչությունը, թվում է, ամենամոտն են օպտիմալ բազմազանությանը: Կենսաբազմազանության որոշ համապատասխան օրինաչափություններ կարելի է գտնել ինչպես գլոբալ, այնպես էլ լանդշաֆտային մասշտաբով:
Բնության կառավարման պրակտիկայի համար դա նշանակում է, որ պահպանման առաջնահերթությունների չափանիշը պետք է լինի մարդածին տեղաշարժի հեռավորությունը օպտիմալ բազմազանությունից, բայց ոչ բարձր ֆորմալ բազմազանության ինդեքսները (օրինակ՝ տեսակների հարստությունը), քանի որ անկայուն կամ սակավ պայմաններին հարմարեցված համայնքներն ունեն համեմատաբար ցածր տեսակների բազմազանություն, որը, այնուամենայնիվ, ապահովում է դրանց առավելագույն գործունեությունը: 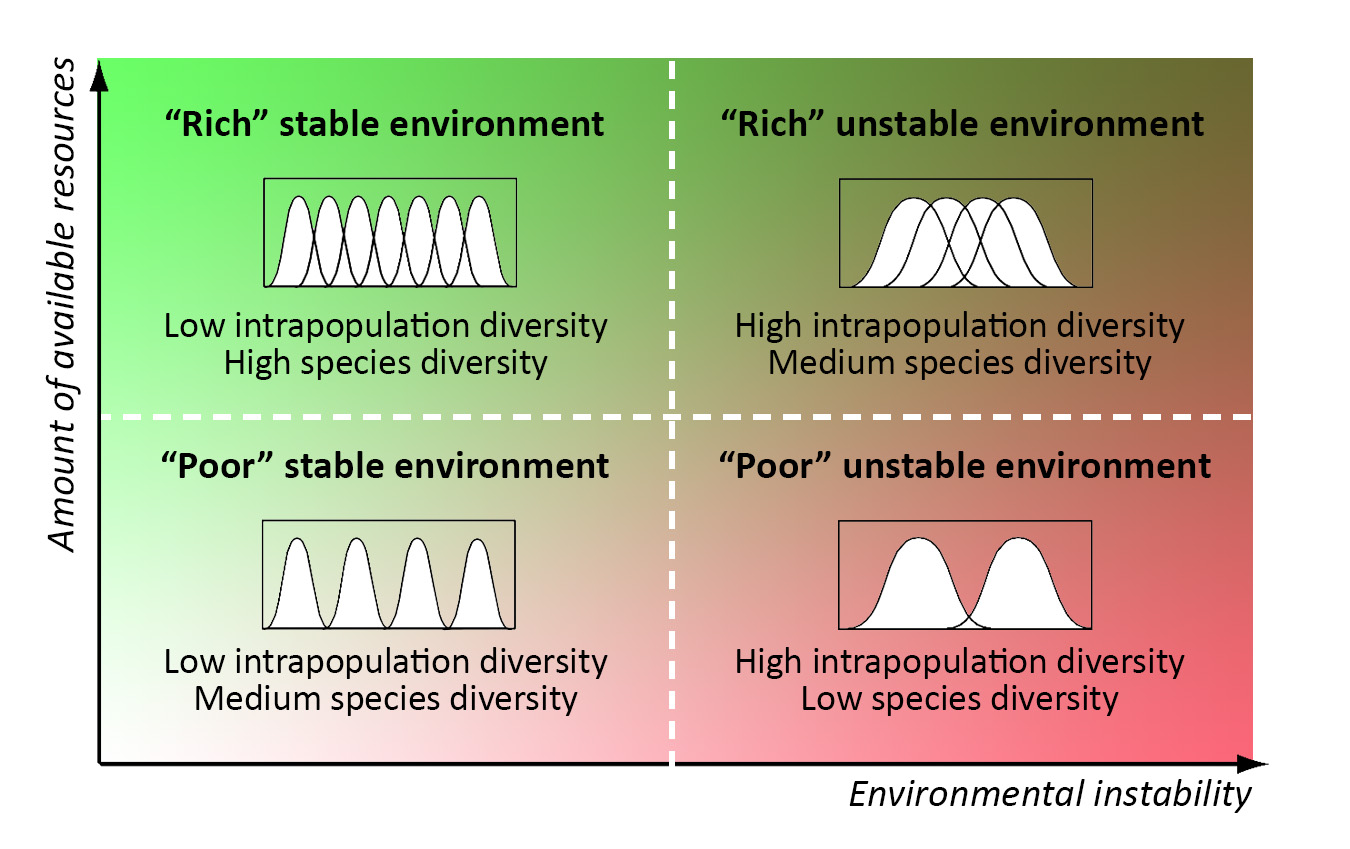
Օպտիմալ տեսակների և ներբնակչության բազմազանության ակնկալվող արժեքները այն համայնքներում, որոնք հարմարեցված են տարբեր միջավայրերին:
Օպտիմալության հավերժական ձգտում: Կարո՞ղ է կենսաբանական համակարգը հասնել օպտիմալ բազմազանության: Դա քիչ հավանական է թվում մեր փոփոխական տիեզերքում: Շրջակա միջավայրի կամ կենսահամակարգերի ցանկացած բնական կամ մարդածին փոփոխություն դրանք իրենց օպտիմալ վիճակից տեղափոխում է ոչ օպտիմալ վիճակի: Այսպիսով, կենսահամակարգերը մշտական «հետապնդման» մեջ են էկոլոգիական, միկրոէվոլյուցիոն և էվոլյուցիոն գործընթացների ժամանակ օպտիմալ վիճակների համար (որոնք կարող են տեղի ունենալ միաժամանակ):
Էկոլոգիական օրինակ 1. r-ստրատեգների գերակշռությունը սինանտրոպ բիոտայում: Շրջակա միջավայրի մարդածին փոփոխությունների ընդհանուր ուղղությունը ապակայունացումն է, մինչդեռ բնակչության վրա ազդեցությունն արտահայտվում է ներբնակչության բազմազանության նվազմամբ: Վերջինս նվազեցնում է բնակչության ապակայունացմանը հարմարվելու հնարավորությունը։ Արդյունքում, ադապտացիան սկսում է գործել համայնքի մակարդակով, և տիպիկ բնիկ տեսակները փոխարինվում են այլ տեսակներով, որոնք փոխում են տեսակների կազմը K-ստրատեգիստներից դեպի r-ռազմավարներ և մասնագետներից՝ ընդհանուր, ինչը համապատասխանում է սինանտրոպ բիոտայի բազմացմանը:
Էկոլոգիական օրինակ 2. համայնքների հավաքման մեխանիզմներ. Որոշ մեխանիզմների տարածվածությունը համայնքի հավաքագրման մեջ որոշվում է «հարստության» և շրջակա միջավայրի կայունության հարաբերակցությամբ, ավելի շուտ(՞), ռեսուրսների որոշակի քանակի ներքո գոյություն ունեցող օպտիմալ նիշերի քանակի և շրջակա միջավայրի պարամետրի (օրինակ՝ t °C) տատանումների (6) տատանումների (6). խորշերը մոտավորապես հավասար են (R/R*≈D/D*); 2) չեզոք մեխանիզմներն աշխատում են հիմնականում շատ «հարուստ» միջավայրերում, որտեղ մեծ քանակությամբ ռեսուրսներ թույլ են տալիս գոյություն ունենալ շատ ավելի մեծ տեսակների քանակ, քան թույլ է տալիս շրջակա միջավայրի պարամետրի շրջանակը (R/R*>> D/D*); 3) «աբիոտիկ ֆիլտրերը» գործում են հիմնականում անպտուղ (կոշտ) միջավայրերում, որտեղ ռեսուրսների փոքր քանակությունը թույլ է տալիս գոյություն ունենալ շատ ավելի փոքր տեսակների քանակ, քան թույլ է տալիս շրջակա միջավայրի պարամետրի միջակայքը (R/R*<< D/D*): 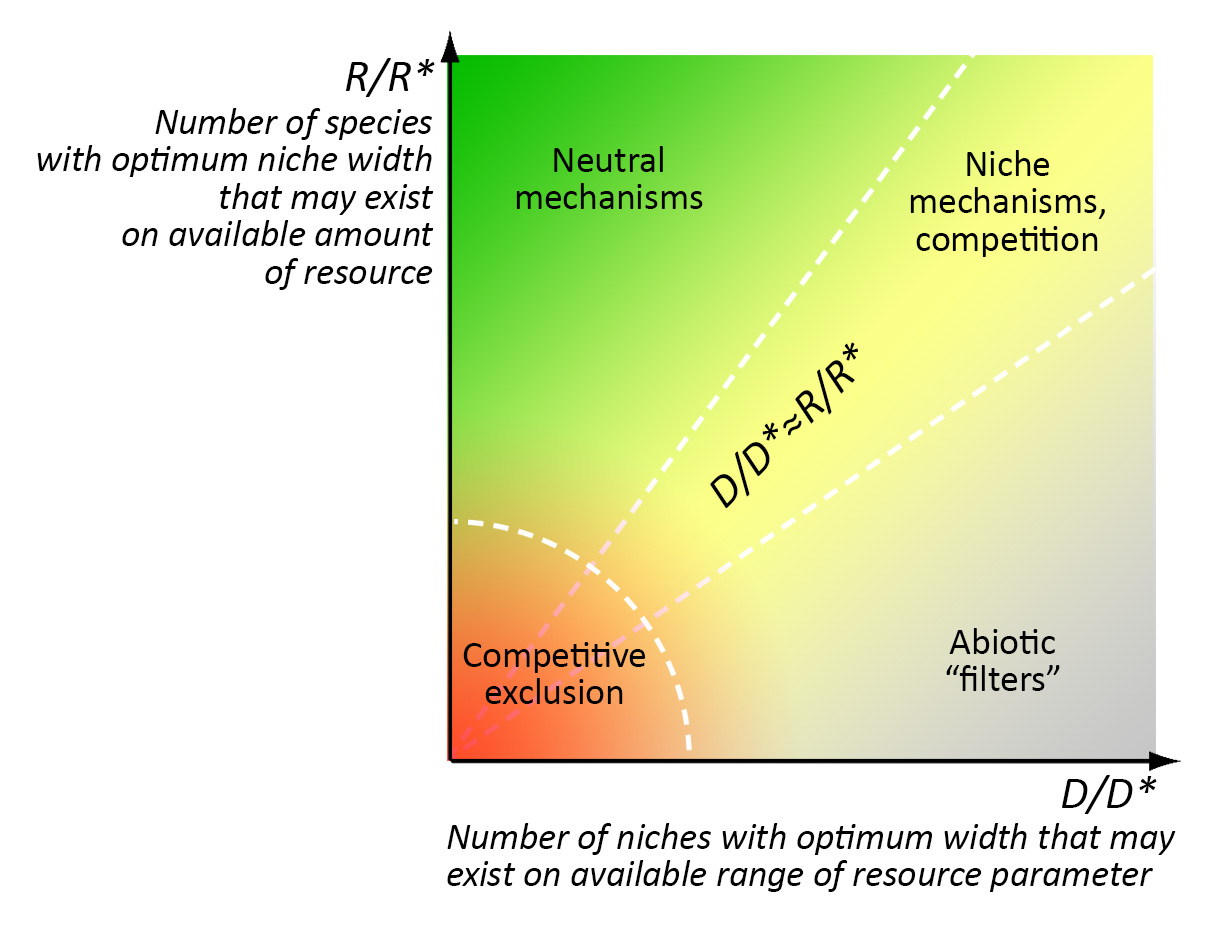
Համայնքների հավաքների տարբեր մեխանիզմների առաջնահերթ գործողության գոտիներ. R-ը հասանելի ռեսուրսների ընդհանուր քանակն է, R*-ը ռեսուրսների քանակն է, որն անհրաժեշտ է օպտիմալ խորշի լայնությամբ բնակչության համար, D-ը ռեսուրսների պարամետրի ընդհանուր միջակայքն է, D*-ը էկոլոգիական խորշի օպտիմալ լայնությունն է:
Միկրոէվոլյուցիոն օրինակ. սիմպատիկ ներտեսակային ձևերը՝ որպես բազմազանության օպտիմալացման մեխանիզմ: Եթե շրջակա միջավայրի կայունության աստիճանը փոխվում է, համայնքը պետք է տեղափոխվի տարբեր օպտիմալ վիճակների միջև՝ կայուն պայմաններում նեղ խորշերով մեծ թվով տեսակներ և անկայուն պայմաններում լայն խորշերով փոքր թվով տեսակներ: Խնդիրներ չկան, եթե մարզային ավազանում կան բավարար տեսակներ, և համայնքը կարող է հեշտությամբ ներառել այնպիսի տեսակներ, որոնք ունեն օպտիմալության հասնելու համար անհրաժեշտ հատկանիշներ: Բայց ի՞նչ, եթե համայնքը մեկուսացված է կամ մարզային ավազանում համապատասխան տեսակներ չկան: Այս դեպքում օպտիմալացում կարող է առաջանալ համակրելի ներտեսակային ձևերի (էկոտիպերի) ձևավորման շնորհիվ, երբ մի տեսակ զբաղեցնում է մի քանի խորշեր: Այս գործընթացը կարող է օպտիմալացնել ինչպես ներտեսակային բազմազանությունը (նիշի լայնությունը), այնպես էլ համայնքի բազմազանությունը (տեսակների թիվը): Սիմպատրիկ ներտեսակային ձևերը կարող են աշխատել որպես դինամիկ համակարգ՝ մշտապես հարմարեցնելով տեղական կենսաբազմազանությունը շրջակա միջավայրի փոփոխություններին, մինչև ձևերի մեկուսացումը հանգեցնի նոր տեսակների ձևավորմանը: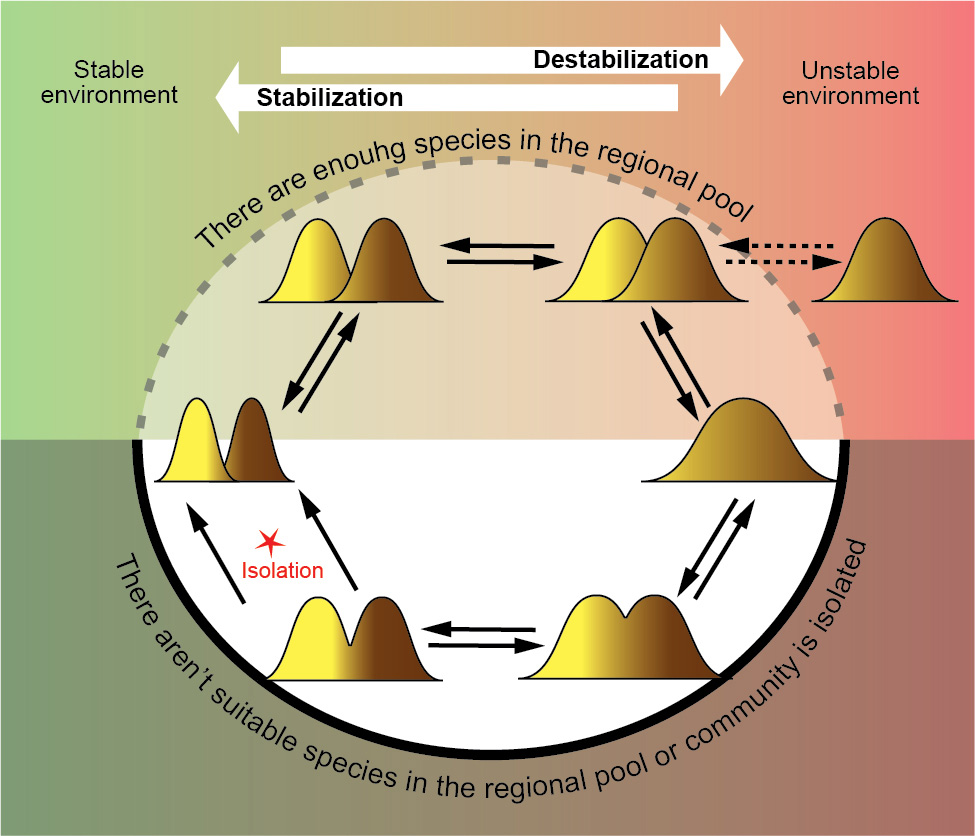
Համայնքների բազմազանության օպտիմալացման դինամիկ ցիկլը. վերին մասը ցույց է տալիս օպտիմալացում, երբ բավարար տեսակներ փոխանակվում են տարածաշրջանային ավազանի հետ, իսկ ստորին մասը ցույց է տալիս օպտիմալացում, երբ համայնքը մեկուսացված է:
Էվոլյուցիոն օրինակ. բազմազանության ընդհանուր աճ՝ չնայած վերելքներին և վայրէջքներին: Կենսաբազմազանության օպտիմալացումը կարող է լրացուցիչ բացատրություն լինել Վ. Ժերիհինի` էվոլյուցիայի բիոցենոտիկ կարգավորման և Վ. Կրասիլովի` համահունչ-անկոհերենտ էվոլյուցիայի հայեցակարգի գործընթացների մեջ: Շրջակա միջավայրի ապակայունացման և համայնքների ոչնչացման պատմական ժամանակաշրջանները (բիոցենոտիկ ճգնաժամ) բնութագրվում են ներբնակչության բազմազանության աճով, խորշերի ընդլայնմամբ, մասնագիտացված տեսակների ոչնչացմամբ, տեսակների բազմազանության անկմամբ: Ներբնակչության բազմազանության աճը նյութ է տալիս ներտեսակային ձևերի և տեսակավորման ապագա ձևավորման համար: Շրջակա միջավայրի կայունացման և համայնքների «նոր սերնդի» զարգացման պատմական ժամանակաշրջանները բնութագրվում են ներբնակչության բազմազանության նվազմամբ, խորշերի նեղացմամբ, ներտեսակային ձևերի դիսկրետացմամբ, տեսակավորման, տեսակների թվի աճով: Օրգանիզմների էվոլյուցիոն մակարդակի աճը հանգեցնում է ներբնակչության բազմազանության նվազմանը (օպտիմալ խորշերը նեղացան) և տեսակների օպտիմալ բազմազանության ավելացմանը (տես վերևում), ինչը կարող է լրացուցիչ բացատրություն լինել կյանքի էվոլյուցիայի ընթացքում տեսակների բազմազանության աճի ընդհանուր միտումի: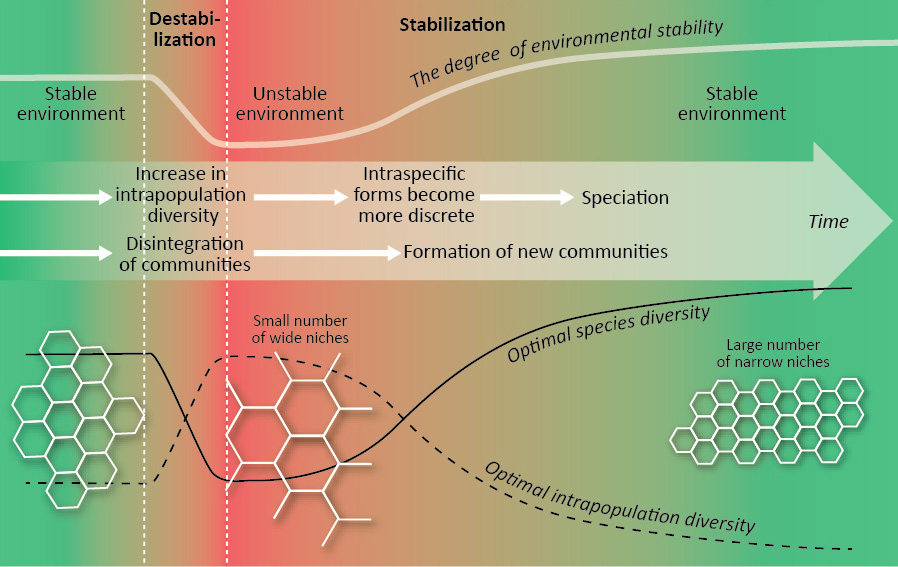
Կենսացենոտիկ ճգնաժամի ընթացքում տեսակների օպտիմալ արժեքների և ներբնակչության բազմազանության փոփոխություններ և նոր համայնքների հետագա զարգացում: Մոխրագույն» բջիջ» ցույց է տալիս էկոլոգիական խորշերի օպտիմալ երկրաչափությունը:
Հիմնական հղումներ
Բուկվարևա, Ե. 2017 թ. Օպտիմալ կենսաբազմազանություն – Լանդշաֆտի գնահատման նոր հարթություն: Էկոլոգիական ցուցանիշներ, 94(2): 6-11. DOI 10.1016/j.ecolind.2017.04.041
Bukvareva E. 2014 թ. Կենսահամակարգերի օպտիմալ բազմազանության սկզբունքի ամփոփում. LAP Lambert Academic Publishing. 52 էջ
Bukvareva E., Aleshchenko G. 2013 թ. Օպտիմալացում, խորշ և չեզոք մեխանիզմներ կենսաբազմազանության ձևավորման մեջ // Life Sciences ամերիկյան ամսագիր, 1 (4): 174-183. DOI 10.11648/j.ajls.20130104.16
Букварева Е.Н., Алещенко Г.М. 2013 թ. Принцип оптимального разнообразия биосистем. М.: КМК-Товарищество научных изданий. 522 с.
Bukvareva E., Aleshchenko G. 2012 թ. Օպտիմալ կենսաբազմազանության և էկոհամակարգի գործունեության սկզբունքը // Էկոհամակարգի միջազգային հանդես, 2 (4): 78-87:
Букварева Е.Н., Алещенко Г.М. 2012 թ. Разделение ниш – условие или следствие наблюдаемого видового разнообразия? Оптимизация разнообразия как дополнительный механизм формирования структуры экологических сообществ //Успехи современной биологии, Т. 132, № 4, с. 337 – 352 թթ.
Ալեշչենկո Գ.Մ. և Բուկվարևա ԵՆ 2010 թ. Օպտիմալ կենսաբանական բազմազանության երկաստիճան հիերարխիկ մոդել // Կենսաբանության տեղեկագիր, 37(1): 1–9.
Букварева Е.Н., Алещенко Г.М. 2010 թ. Оптимизация разнообразия надорганизменных систем как один из механизмов их развития в экологическом, микроэволюционном и эволюционном масштабах // Успехивременной биологии. Т.130. № 2. С. 115-129 թթ.
Алещенко Г.М., Букварева Е.Н. 2010 թ. Двухуровневая иерархическая модель оптимизации биологического разнообразия // Известия РАН, серия биологическая, №1, с. 5-15